
La présence en Guyane d’Hydrilla verticillata (L.f.) Royle, plante immergée de la famille des Hydrocharitacées, est maintenant validée.
La découverte de l’espèce a été signalée dans une actualité de « Sentinelles de la Nature » du 15 avril 2025 et par deux observations insérées sur le réseau iNaturalist datées d’aout 2025, l’une la positionnant dans le même plan d’eau du quartier Hibiscus à Cayenne, avec une population en fleurs, l’autre dans le Morne Coco dans la commune de Remire-Montjoly avec une population dépourvue de fleurs (« toujours stérile 6 ans après » une première observation, selon le découvreur).
Le premier site, très fortement colonisé par la plante, a fait l’objet d’un curage durant l’été 2025. Réalisée par une entreprise privée pour le compte de la Communauté d’Agglomération du Centre Littoral (CACL), cette intervention d’une durée de 3 semaines a montré une assez faible efficacité car un nouveau développement de l’espèce a été assez rapidement observé dans le plan d’eau.
Par ailleurs, lors d’un contact très récent avec Thibault Lebrun de l’association Kwata, il a signalé la découverte le 27 mars 2026 d’une mare colonisée par Hydrilla verticillata dans la zone des Salines de Montjoly. Après un arrachage des plantes le 3 avril, la mare a été comblée le 8 avril. Un suivi hebdomadaire du site a été mis en place pour vérifier l’absence de nouvelles repousses. Une actualité signalant deux alertes successives d’espèces exotique envahissantes dans ces Salines, dont celle portant sur hydrilla, a été mise en ligne le 26 avril sur le site de l’association.
Enfin, un accès le 23 juin au réseau INaturalist a permis de constater une augmentation notable des observations insérées sur l’espèce en Guyane (voir la carte), avec au total près d’une quinzaine de signalements dans le secteur du plan d’eau du quartier Hibiscus dont quelques uns en aval, une deuxième observation dans les Salines de Montjoly et six signalements dans le secteur de Morne Coco.
Si ces observations se trouvent dans ou à proximité de zones urbanisées, rien de vraiment surprenant, compte tenu de l’attrait esthétique de l’espèce pour les aquariophiles, en revanche une autre, datant du 24 avril, située à une trentaine de kilomètres dans le Parc Naturel de Guyane dans la crique Gabriel, un site beaucoup moins fréquenté, est nettement distante de toutes les autres.
A l’heure actuelle, aucune prospection systématique concernant cette espèce n’a encore été menée sur le territoire guyanais.
Le présent article est une alerte destinée à tous les publics et personnes concernées à divers titres par les dommages éventuels que pourrait causer la dispersion de cette espèce à la biodiversité et aux usages humains des milieux aquatiques d’eau douce de la Guyane.
En effet, présentée comme « The Perfect Aquatic Weed » par Kenneth A. Langeland dans une publication de 1996, pour sa dispersion rapide aux Etats Unis et les nombreuses nuisances qu’elle y causait, la multiplication des implantations de cette espèce sur de très nombreux territoires durant ces dernières décennies en a fait une des plantes aquatiques exotiques les plus suivies et étudiées à l’échelle mondiale.
Description
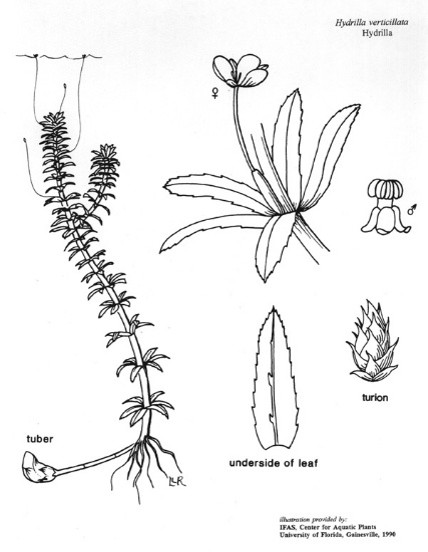
Cette plante est constituée de longues tiges ramifiées pouvant atteindre 9 m de longueur et former des tapis très denses vers la surface. Elles peuvent développer des stolons au contact du substrat et des rhizomes enfouis jusqu’à 30 cm dans le sédiment (Langeland, 1996), des structures pouvant participer à l’étalement progressif de la plante. Des racines adventives simples peuvent être présentes aux nœuds des tiges.
A la base des tiges peuvent se trouver des feuilles opposées dépassant rarement 4 mm de longueur mais sur toute leur longueur elles portent des verticilles composés généralement de 3 à 8 feuilles se terminant en pointe et mesurant de 2 à 4 mm de large sur 6 à 20 mm de long (Cook & Lüönd, 1982). Les bordures de ces feuilles présentent une dentelure visible à l’œil nu et des dents (ou épines) peuvent également se trouver sur la nervure principale de leur face inférieure.
N. B. : Cette dentelure visible est la caractéristique permettant le plus facilement d’identifier cette espèce parmi les autres espèces de la famille des Hydrocharitacées aux morphologies assez proches, telles que Elodea spp., Egeria spp. ou Lagarosiphon spp.. La présence de verticille (absence chez Lagarosiphon spp) et le nombre de feuilles par verticille (trois seulement chez Elodea spp., 4 ou 5 chez Egeria spp.) sont des critères de discrimination également utilisables.

L’espèce peut présenter deux types de formes, l’une dioïque, aux pieds mâles et femelles séparés, et une forme monoïque aux organes mâles et femelles sur les mêmes plantes.
Ses inflorescences sont installées vers l’extrémité des tiges.
Portées par un pédoncule pouvant atteindre 10 cm, les fleurs femelles flottant en surface comportent trois sépales blanchâtres et trois pétales translucides de 10 à 50 mm de long et de 4 à 8 mm de large.
Présentant trois sépales rouge blanchâtre ou brun de 3 mm de long et 2 mm de large et trois étamines, les fleurs mâles, immergées sur un pédoncule court, se détachent des tiges à maturité et, en flottant sur l’eau, peuvent venir polliniser les fleurs femelles (Cook & Lüönd, 1982 ; Langeland, 1996 ; Yeo et al., 1984).

N.B. : pour contribuer à son identification, un accès à la fiche consacrée à l’espèce sur le site iNaturalist rassemblant des observations mondiales permet de visualiser de très nombreux clichés de la plante pouvant faciliter sa reconnaissance.
Des particularités biologiques et écologiques
La plupart des types de milieux aquatiques d’eau douce stagnants ou à faible courant, naturels ou artificiels, peuvent accueillir des populations d’H. verticillata : lacs, étangs, zones humides peu profondes, retenues, bassins urbains, canaux, cours lentiques des fleuves et rivières, etc. Elle peut également se rencontrer dans des milieux saumâtres ayant une salinité pouvant aller jusqu’à 7 ‰ (True-Meadows et al., 2016), avec un seuil de dommage mesuré en laboratoire de 13 ‰ (Steward & Van, 1987).
Ses populations colonisent généralement des zones peu profondes, ne dépassant pas 3 mètres environ, mais dans des milieux aux eaux transparentes elle est susceptible de s’installer jusqu’à 15 mètres de profondeur (Yeo et al., 1984). L’espèce peut s’installer dans des conditions de lumières très faibles par rapport à d’autres espèces de plantes immergées (Van et al., 1976). Avec une photosynthèse encore active jusqu’à des intensités lumineuses très faibles, de 10 à 12 lmol m⁻² s⁻¹ selon Bowes et al. (1977), elle peut aussi prospérer dans des eaux relativement troubles, comme par exemple celles du fleuve Potomac, aux États-Unis, avec une transparence mesurée au disque de Secchi d’environ 0,60 m (Carter & Rybicki, 1986).
Ses très larges capacités d’adaptations à de multiples conditions environnementales se manifestent également par l’amplitude thermique tolérée puisque l’espèce est capable de subsister dans des eaux depuis 10 à 15°C jusqu’à au moins 30°C, avec un optimum thermique entre 20 et 27°C, et dans des qualités des eaux très variables en termes de pH (5,7 à 8,5) et de teneurs en nutriments (Gu, 2006).
Les biomasses maximales mesurées dans des développements denses de l’espèce dans différents plans d’eau aux Etats-Unis dépassent 900 g/m² de matières sèches (Haller & Sutton, 1975 ; Bowes et al., 1979 ; Shields et al., 2012). Ces valeurs sont assez proches des maximas mesurés pour d’autres plantes immergées, mais dans des travaux menés sur le fleuve Paraná au Brésil, Sousa et al. (2009) citent une valeur dépassant 1400 g/m².
Hormis ce dernier chiffre, telles que présentées jusqu’ici, les caractéristiques biologiques et écologiques de l’espèce ne font pas d’Hydrilla verticillata une espèce apparemment exceptionnelle en tant que phanérogame immergée des eaux douces, ni en diversité de milieux aquatiques occupés, de gamme de profondeur, de qualité d’eau ou de production végétale.
Une reproduction végétative particulièrement efficace
Ses caractéristiques permettent en effet, et sans hésitation, d’attribuer à l’espèce un caractère tout à fait remarquable – sans aller jusqu’à celui de perfection cité par Kenneth A. Langeland – et ainsi de pleinement justifier la présente alerte.
Rappelons que pour de très nombreuses plantes aquatiques immergées, la reproduction sexuée faisant intervenir floraison, pollinisation puis développement de graines viables, rencontre souvent d’importantes contraintes induites par les conditions environnementales locales, ce qui conduit à une reproduction strictement végétative par un développement continu des tiges de ces espèces et la production de stolons et/ou de rhizomes.
Pouvant s’installer à des profondeurs très variables, H. verticillata produit depuis le substrat des tiges souvent peu ramifiées dont l’architecture se modifie à proximité de la surface par une multiplication des ramifications, amenant à une sorte de canopée pouvant occuper tout le volume dans les derniers 50 cm des eaux, correspondant alors à environ 50 % de la biomasse de la plante (Gu, 2006).
Ce tapis de surface permet à la plante d’accéder à toute la lumière disponible pour en faciliter l’activité photosynthétique et peut ainsi induire une concurrence gagnante sur les autres espèces immergées présentes. Par exemple, Haller & Sutton (1975) ont observé qu’une réduction de la pénétration de la lumière d’au moins 95 % pouvait se produire dans les 30 premiers centimètres d’une telle canopée dense, privant de lumière les autres plantes immergées présentes dans le biotope.
Des tiges très dynamiques
Ces développements rapides des tiges créant des herbiers denses dans de nombreux sites ont bien sûr été l’objet de recherches. Par exemple, Glomski & Netherland (2012) ont répondu à la question débutant le titre de leur article « Does hydrilla grow an inch per day? » par des travaux en mésocosmes comparant durant cinq semaines les productions de tiges latérales, de nouvelles tiges et de stolons d’hydrilla avec trois autres plantes immergées, dont le myriophylle en épi (Myriophyllum spicatum), exotique en Amérique, et deux espèces américaines, l’élodée du Canada (Elodea canadensis) et l’Hétéranthère litigieuse (Heteranthera dubia).
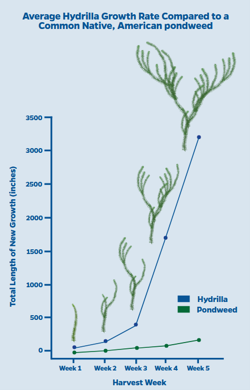
Dans ces conditions expérimentales, durant cinq semaines en période estivale, hydrilla a pu produire plus de 150 tiges latérales, 190 tiges nouvelles et 35 stolons alors que les autres espèces ont montré de très faibles développements, dont aucun stolon et au maximum une douzaine de tiges latérales et huit tiges nouvelles. A partir d’une tige de 10 cm de longueur d’hydrilla, la longueur cumulée des tiges et stolons en fin d’expérimentation était supérieure à 8 mètres.
Ces résultats ont d’ailleurs conduit le « Center For Aquatic And Invasive Plants » de l’Université de Floride à diffuser une plaquette sur ces résultats pour illustrer l’impressionnante vitalité de la plante.
Comme pour d’autres plantes de conformation similaire, les tiges fines et longues de l’hydrilla peuvent se briser assez facilement sous l’effet de perturbations mécaniques (action des vagues et des courants, activités humaines, animaux, etc.) pour être ensuite dispersées dans le milieu. Dans des conditions favorables, ces fragments, même de très petite taille, peuvent alors constituer autant de propagules ou boutures viables, capables de subsister des jours sinon des semaines dans les eaux pour être transportées sur des distances variables selon les milieux.
Les capacités de repousses de fragments de tiges à partir des turions ont été évaluées par Langeland & Sutton (1978) en laboratoire et en mésocosmes. Les tests d’une durée d’un mois ont porté sur des fragments de tiges comportant de 1 à 5 nœuds.
Pour ces deux conditions expérimentales, des repousses ont été observées dans toutes modalités, soit dans 28 % des fragments ne comportant qu’un seul nœud jusqu’à 98 % (mésocosme) et 100 % (laboratoire) pour les fragments à cinq nœuds. Les augmentations de poids (repousses) des fragments étaient proportionnelles au nombre de nœuds. Une démonstration supplémentaire de la vitalité de l’espèce !
Dans une expérimentation en mésocosme d’un an et demi, Madsen & Smith (1999) ont par ailleurs cherché à évaluer les capacités relatives de la production de fragments et de stolons dans la propagation latérale de la plante à partir d’individus plantés. Dans ces conditions expérimentales en l’absence de courant, les quelques fragments produits n’ont permis que 0,1 % d’implantations nouvelles de plantes enracinées. Les 99,9 % d’implantations nouvelles par les stolons ont montré des allongements pouvant atteindre 4,0 cm j⁻¹ ce qui est relativement important.
Hydrilla a fait partie des 19 espèces de plantes aquatiques dont des fragments ou des individus dérivants ont été récupérés en 2000 dans la rivière San Marcos au Texas lors de prélèvements sur plusieurs sites aux quatre saisons (Owens et al., 2001). Les fragments de tige d’H. verticillata ont été placés en mésocosmes durant un mois avant d’être examinés. Il s’agissait d’évaluer les capacités d’établissement de ces fragments en vérifiant leur production de racines : 70 % (juin) et 83 % (mars) de ces fragments s’étaient enracinés après un mois de croissance et donc considérés comme « établis ».
Enfin, malgré la relative fragilité de ces tiges, il semble qu’elles puissent résister à un certain dessèchement car Sousa (2011) cite des travaux de Silveira et al. (2009) dans lesquels des fragments d’extrémité de tiges d’Hydrilla étaient capables de se régénérer même après 4 jours d’exposition à l’air sur un sédiment argileux.
Des fragments comportant un ou quatre verticilles desséchés à 26 °C pendant 0 à 8 heures ont fait l’objet d’un suivi pendant 14 jours après leur réintroduction dans l’eau par Baniszewski et al. (2016). Les fragments à un verticille desséchés pendant 2 heures présentaient un faible taux de survie après dessiccation par rapport aux fragments à quatre verticilles mais pour les durées de dessèchement plus longues ces taux étaient proches de zéro pour tous les fragments. De même, les durées longues de dessèchement réduisaient fortement les germinations des fragments par rapport aux témoins.
Enfin, des parties de l’extrémité des tiges soumises à un séchage à 30 °C pendant 16 H dans une humidité relative de 40 % étaient encore physiologiquement actives selon Basiouny et al. (1978).
Tubercules et turions : des moteurs cachés de dispersion
En matière de capacité de développement végétatif, cette importante vitalité des tiges de la plante est très largement complétée par sa capacité de produire deux types d’organes végétatifs permettant à l’espèce de résister à des conditions défavorables, en particulier en hiver, d’où le nom d’hibernacles qui peut leur être attribué. Il s’agit de tubercules se formant aux extrémités des stolons et des rhizomes dans les sédiments, et de turions sur les tiges. Les tubercules sont également présentés comme des turions souterrains (« subterranean turions« ).
Les tubercules de 5 à 10 mm de longueur sont des organes de réserve pouvant produire jusqu’à 5 000 à 6 000 nouveaux tubercules par m². Les turions sont des bourgeons foliaires vert foncé, compacts et pointus, de 5 à 8 mm de long, se développant à l’aisselle des feuilles, tombant de la plante à maturité comme de nouvelles boutures. Chaque turion peut produire jusqu’à 3 000 nouveaux turions par m² (Langeland, 1996).
Ces valeurs élevées ont été généralement obtenues lors d’expérimentations en mésocosme. Dans l’article de True-Meadows et al. (2016) sont rassemblées des données issues de prélèvements en milieux naturels proposant une gamme de densités plus faibles, de moins de 100 à environ 1700 tubercules par m². Par exemple, dans les trois lacs de Floride étudiés par Bowes et al. (1979), ces valeurs ne dépassaient guère 500 tubercules par m².

Des tests destinés à évaluer la durée de vie fonctionnelle de ces organes ont été réalisés sur 4 ans par Van & Steward (1990). Une vérification de l’état des tubercules et des turions utilisés dans ces expérimentations en mésocosmes avait montré des taux de germination initiaux de respectivement 94 et 97 %. Si aucun turion ne durait plus d’une année, les tubercules présentaient encore des germinations, moins nombreuses, au bout de la deuxième année pour n’en produire aucune la quatrième année. Dans la discussion de leur article, les auteurs suggéraient, tout comme l’avaient déjà fait Spencer et al. (1987), que les différences de durée de vie dede ces deux types d’organes pouvaient constituer des mécanismes successifs complémentaires améliorant encore les capacités de développement de l’espèce. C’est également ce que montrent les tests de Basiouny et al. (1978) avec des tubercules nettement plus résistants au dessèchement que les turions, conservant un taux de germination de 16,7 % après 64 h, alors que ce taux pour les turions diminue très fortement en 8 heures.
Un commentaire

Dans leur ouvrage « Hydrilla Integrated Management« , daté de 2014 mais revu en 2020, Gillet-Kaufmann et ses collègues résument les capacités colonisatrices de l’espèce en citant la tolérance à différents habitats, l’absence d’ennemis naturels aux USA, une croissance rapide, la concurrence pour la lumière et la propagation par turions, tubercules et fragments des tiges. La vitalité des tiges de la plante et ses productions de tubercules et turions jouent donc un rôle très important dans ses capacités de colonisation.
Dans son article, Langeland (1996) signale même que selon une publication de Joyce et al. (1980) les tubercules pourraient survivre après ingestion et régurgitation par les oiseaux d’eau.
Rien de surprenant car des recherches sur ce point ont montré la possibilité de consommation par des oiseaux de graines et de propagules restant ensuite viables, dans des ouvrages anciens comme ceux de Sculthorpe (1967) ou Van der Pijl (1969), ou plus récemment de Lovas-Kiss et al., 2018).
N. B.: cette publication « Joyce, J. C., W. T. Haller and D. E. Colle. 1980. Investigation of the presence and survivability of hydrilla propagules in waterfowl. Aquatics 2:10-14. » n’a pu être consultée.
Répartition
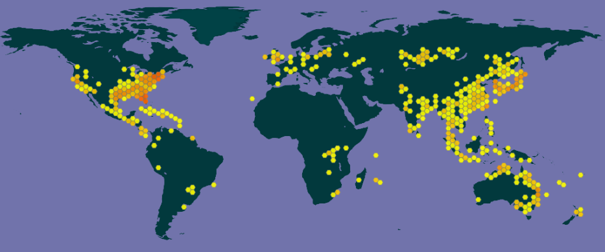
La carte du GBIF (Système mondial d’information sur la biodiversité) qui est consacrée à cette espèce montre l’ampleur de sa répartition planétaire actuelle.
Aire d’origine de l’espèce
Selon Cook & Lüönd (1982), elle se situerait « dans les régions les plus chaudes d’Asie ». Les recherches plus récentes indiquent qu’elle pourrait couvrir une plus large zone géographique allant du sud de l’Asie au nord de l’Australie mais il reste très difficile de la définir précisément car nombre des informations disponibles restent vagues : par exemple selon une base de données européenne certaines des populations observées en Europe seraient d’origine incertaine « doubtfully native »…
Cependant, grâce aux analyses génétiques, il semblerait acquis que sa forme monoïque soit originaire de Corée et les plantes femelles de la forme dioïque du sud de l’Inde (Madeira et al., 1997), et la forme dioïque mâle du nord de l’Australie (Hofstra et al., 2000).
Avec le développement des analyses génétiques, une publication plus récente (Tippery et al., 2020) signalait l’existence d’une nouvelle souche d’Hydrilla verticillata introduite dans la rivière Connecticut au Nord-Est des Etats Unis, probablement originaire du nord de l’Eurasie. Plus récemment encore, les travaux de Tippery (2023) ont conduit à identifier cinq principaux biotypes de l’espèce ce qui permet à l’auteur de proposer une clé de détermination morphologique de cinq sous-espèces (en Annexe), pour améliorer la communication mondiale sur cette espèce aussi problématique.
Dispersion
La dispersion de l’espèce sur la planète n’a rien de surprenant car, très appréciée comme plante aquatique d’ornement en aquarium, elle a fait depuis des décennies l’objet d’échanges commerciaux mondiaux (un exemple actuel). Le rejet ou l’abandon de fragments de l’espèce en milieu naturel par des aquariophiles négligents et/ou ignorants, voire des implantations réalisées pour un intérêt esthétique, expliquent probablement toutes les multiples installations constatées dans de nouveaux territoires.
Cette dispersion intercontinentale via les échanges commerciaux d’aquariophilie explique que des plantes des trois formes qui se ressemblent fortement, sauf durant la période de floraison, aient pu être transportées séparément et coloniser ainsi des territoires particuliers. C’est ainsi, par exemple, que dans leur article identifiant la provenance depuis la Malaisie-Indonésie de l’hydrilla introduite en Afrique du Sud, Madeira et al. (2007) présentaient une arborescence de la génétique des biotypes de 42 populations d’hydrilla introduites dans divers pays en y précisant les localisations géographiques des trois biotypes.
Compte tenu des possibilités de survie déjà décrites des diverses propagules de l’espèce (tiges, tubercules et turions), leur transport involontaire à une échelle régionale ou locale lors de déplacements de quelques jours entre milieux aquatiques par des usagers ou des entreprises avec leur matériel peut permettre une arrivée et une éventuelle installation de l’espèce dans un nouveau site. Ces propagules clandestines peuvent également être transportées par des animaux lors de leurs déplacements, par exemple par des oiseaux (Coley, 1997).
Ces propagules peuvent aussi se retrouver comme « passagères clandestines » d’autres plantes commercialisées comme ornement : dans la synthèse sur l’espèce en Nouvelle-Calédonie (Flouhr & Mary, 2010), un exemple était cité à propos de l’arrivée en 1994 d’hydrilla dans deux lacs proches de Seattle (état de Washington), Pipe et Lucerne, où l’introduction était attribuée soit à une vidange d’aquarium soit à une de ces clandestines apportée avec des nénuphars exotiques… L’installation et la colonisation très importante de ces deux lacs par hydrilla avait rapidement conduit à des interventions pour l’éradiquer. De tels exemples de « passagères clandestines » avaient aussi été signalés dans les résultats d’une enquête menée en Belgique sur le commerce des plantes aquatiques (Van den Neucker & Scheers, 2022).
A ces échelles géographiques, les processus de dispersion de proche en proche de longues durées peuvent tout à fait s’exercer en fonction d’activités régulières ou saisonnières (pêche, loisirs nautiques, etc.) entre milieux aquatiques voisins, comme cela a pu être par exemple observé pour d’autres plantes exotiques dans les lacs et étangs du littoral aquitain (Ribaudo et al., 2022)
Répartition en France
Depuis quelques années, des fragments de cette espèce ont été observés de manière éphémère à quelques reprises dans le Sud-Ouest de l’hexagone mais, à notre connaissance, aucune population installée n’a encore été recensée.
Citée dans la publication de Cook & Lüönd (1982), H. verticillata présente à l’île de La Réunion pourrait y être indigène.
A la Guadeloupe, après une première découverte dans un étang de l’île Marie-Galante en 1991, les travaux de Maddy (2008) ont permis de l’observer dans quatre cours d’eau permanents de la Basse-Terre et quatre étangs dont deux à Marie-Galante. L’Etang Roland sur Basse-Terre (plus de deux hectares) était totalement colonisé par hydrilla. Maddi (2008) terminait son article en rappelant les risques d’invasion de l’espèce dans les milieux aquatiques de l’île. A notre connaissance aucune intervention de gestion n’a été entreprise sur ces populations.
A la Martinique, repérée dans deux mares ornementales en 2008 (Maddi & Brizard, 2010), l’espèce ne semblait pas avoir colonisé d’autres milieux lors des prospections menées en 2013-2014, ce qui avait conduit Maddi (2014) à noter qu’elle demeurait encore très rare. En 2016, la découverte de nouvelles stations de l’espèce dans quatre communes a cependant amené à réaliser une intervention expérimentale d’arrachage manuel de la plante dans un des sites (Delnatte et al., 2021).
Introduite en Nouvelle-Calédonie, après une première observation en 1966 dans une mare artificielle à Nouméa, l’espèce s’est dispersée dans diverses stations en cours d’eau et en plans d’eau (Flouhr & Mary, 2010).
Dans le rapport de l’IRD « Les espèces exotiques envahissantes de Nouvelle-Calédonie » de Hequet et al. (2009), les auteurs précisent que deux espèces au « caractère envahissant évident » évaluées et ajoutées dans la liste prioritaire (Egeria densa et Hydrilla verticillata) n’ont pas été cartographiées par manque d’information et de prospection systématique des cours d’eau et points d’eau du territoire.
Le protocole d’évaluation et de classement des espèces utilisé dans ce rapport était celui développé par NatureServe (Morse et al., 2004) qui permet d’évaluer les taxa pour une région spécifique et d’attribuer à chaque espèce un « rang d’impact » négatif (I-Rank) sur la biodiversité de cette région. Avec un score I-Rank de 38, H. verticillata est en 22ième place du classement de la centaine d’espèces prioritaires et notée « A étudier » dans le tableau récapitulatif des objectifs de gestion.
Une carte de répartition est disponible dans la synthèse bibliographique de Flouhr & Mary (2010) mais les autrices indiquent que ces informations établies à la suite d’une exploitation des bases de données existantes ne permettaient pas d’établir une carte exhaustive.
Une fiche de présentation de l’espèce figure dans le guide « Plantes envahissantes pour les milieux naturels de Nouvelle-Calédonie » (Groupe espèces envahissantes, 2011), pages 88-89.
Elle est également présente dans diverses îles en Polynésie française où elle ne semble pas causer de difficultés particulières.
Impacts et évaluation de risques
Les impacts et les dommages que peuvent créer les développements d’H. verticillata dans les écosystèmes d’accueil s’exercent à la fois sur les communautés végétales et animales et les usages humains des milieux ainsi colonisés. Ils sont de même nature que ceux produits par d’autres plantes immergées mais leur ampleur dépend de leur rapidité de production de biomasse et du degré d’occupation des biotopes. De ce point de vue, H. verticillata semble relativement exceptionnelle.
Des évaluations de ces impacts figurent dans certaines des nombreuses recherches menées sur l’espèce, en particulier en Floride. La fiche présentant l’espèce sur le site « Nonindigenous Aquatic Species » de l’U.S. Geological Survey permet par exemple d’accéder à des informations sur ses divers impacts écologiques, économiques et vis-à-vis de la santé humaine en s’appuyant sur ces références scientifiques disponibles.
Une évaluation des risques liés à H. verticillata a été publiée en juin 2020 par le Service d’inspection sanitaire des animaux et des végétaux du Département de l’Agriculture des États-Unis. Cette première version rassemble informations, références scientifiques utilisées pour cette évaluation et propose un résumé de cette évaluation intitulée « Impact potentiel » :
« L’Hydrilla verticillata figure parmi les pires plantes aquatiques envahissantes au monde (Hofstra & Clayton, 2014), et les responsables américains dépensent chaque année des millions de dollars pour lutter contre cette espèce dans les écosystèmes naturels, les canaux, les réservoirs et les canaux d’irrigation (Manuel et al., 2013 ; Mullin et al., 2000 ; Stocker & Hagstom, 1986). Elle fait de l’ombre et évince les plantes indigènes, car ses tapis denses bloquent la lumière du soleil pour la végétation submergée (Hofstra & Clayton, 2014 ; Langeland, 1996). Plusieurs espèces de poissons ont une taille réduite dans les eaux infestées par H. verticillata (Colle & Shireman, 1980), et les eaux infestées abritent également des populations plus faibles de gastéropodes (Colon-Gaud et al., 2004). Elle est présente dans tous les bassins versants et dans 43 % des lacs publics de Floride (Langeland, 1996), et est donc susceptible d’affecter des écorégions d’importance mondiale (Ricketts et al., 1999). La navigation de plaisance, la baignade et la pêche peuvent devenir difficiles, voire impossibles, dans les eaux infestées (Ramey, 2001 ; Yeo et al., 1984). La plante obstrue les canaux, les stations de pompage et les canaux d’irrigation et affecte la production d’énergie hydroélectrique (Ramey, 2001 ; Sousa, 2011). Au cours des années 1970, elle a affecté 500 miles de canaux d’irrigation en Californie (Mullin et al., 2000). Nous avons évalué le niveau d’incertitude comme très faible pour cet élément de risque en raison de la documentation exhaustive sur l’impact de cette espèce. » (Traduction en ligne DeepL)
N.B. : un risque écologique, voire de santé animale et humaine, assez particulier, est lié au fait qu’H. verticillata peut héberger une cyanobactérie épiphyte, Aetokthonos hydrillicola. Cette cyanobactérie produit une neurotoxine responsable de la myélinopathie vacuolaire aviaire (AVM), une maladie neurologique qui touche depuis le début des années 2000 divers oiseaux aquatiques et leurs prédateurs dans le sud-est des États-Unis, y compris les pygargues à tête blanche (Wilde et al., 2005). Les recherches plus récentes, dont celles de Breinlinger et al. (2021) « Hunting the eagle killer » ou Schwark M. et al. (2023) « More than just an eagle killer« , signalent la toxicité élevée des dérivés de cette toxine induisant une menace potentielle pour la santé humaine.
Statuts attribués à l’espèce. Règlementation en France.
La fiche du GBIF (Système mondial d’information sur la biodiversité) sur l’espèce liste les 33 états ou îles où elle fait l’objet d’un classement dans le registre mondial des espèces introduites et envahissantes. S’y retrouvent la Nouvelle-Calédonie où elle est classée à un niveau de priorité 3 parmi les 70 EEE « établies prioritaires » répertoriées dans la Stratégie de lutte contre les EEE en Nouvelle-Calédonie mise en place en 2016 (CEN, 2016) et la Martinique (OFB, 2021) où elle fait partie des espèces interdites de toutes activités de la famille des Hydrocharitacées, (toutes les espèces sauf Limnobium laevigatum). Bien que non référencée dans la liste du GBIF, la Guadeloupe intègre également une réglementation sur cette espèce.
La règlementation s’appliquant à l’espèce en France hexagonale et en outremer (Guadeloupe, Martinique et Nouvelle-Calédonie, etc.) est rassemblée dans le panorama édité en 2024 par le Centre de ressources Espèces Exotiques Envahissantes. Bien que citée comme présente en Polynésie française, H. verticillata ne fait pas partie des espèces végétales y menaçant la biodiversité (Arrêté de 2006 où se trouve listée Egeria densa, espèce très proche).
Enfin, H. verticillata a été insérée en 2012 dans la liste « of invasive alien plants » de l’Organisation Européenne et Méditerranéenne pour la Protection des Plantes. La fiche qui lui est consacrée indique que bien qu’en Europe sa répartition soit encore limitée, son comportement envahissant sur tous les continents a conduit à la considérer comme une espèce envahissante émergente en Europe.
Quelle stratégie de gestion à appliquer en Guyane ?
Un premier objectif d’une stratégie à mettre en œuvre suite à la découverte d’une nouvelle plante aquatique exotique envahissante est, pour préciser la situation la concernant, de procéder à des prospections systématiques dans les biotopes favorables, dans un premier temps aux alentours des sites colonisés et dans les milieux connectés, puis élargir le périmètre des prospections. L’observation récente dans la crique Gabriel, éloignée de toutes les autres, peut tout à fait être considérée comme un avertissement supplémentaire à une large vigilance. L’identification de l’espèce étant validée, une démarche active de prospection portée par tous les partenaires engagés dans la préservation de la biodiversité en Guyane devrait être lancée dès que possible.
En matière d’interventions de gestion, il semble que les travaux réalisés dans le plan d’eau du quartier Hibiscus aient montré une durée d’efficacité réduite, il y aurait à renouveler l’opération en tenant effectivement compte des caractéristiques biologiques et écologiques de l’espèce pour espérer l’éradiquer du plan d’eau. Les connaissances disponibles sur H. verticillata sont largement suffisantes pour orienter les choix techniques d’intervention (par exemple, en présence ou non de tubercules, intérêt d’un décapage des sédiments). Il semble également nécessaire d’intégrer dès que possible dans les réflexions à mener en matière de stratégie d’actions la diversité des sites où l’espèce est maintenant identifiée pour en examiner spécifiquement les possibilités d’intervention.
Si des informations sur l’ensemble des possibilités de gestion de l’espèce, incluant le recours aux herbicides et/ou à des agents de lutte biologique, sont très largement disponibles (Gillett-Kaufman et al., 2014), les techniques applicables dans la situation actuelle semblent devoir se référer à des interventions mécaniques appropriées, aux conditions pratiques à préciser, ou manuelles si la population à retirer est récente et peu importante. Un arrachage manuel conduit avec précaution par des personnes convenablement formées peut être très efficace.
Rappelons que depuis 2015, des informations sur la gestion des espèces exotiques envahissantes rassemblées à l’échelle nationale en quatre volumes sont disponibles en ligne. Le premier d’entre eux (Sarat et al., 2015) présente les connaissances pratiques sur la gestion, les autres des exemples d’intervention par espèce.
Enfin, ces techniques d’intervention manuelles et mécaniques vont extraire les plantes des sites, aussi les interventions doivent-elles nécessairement prendre en compte le devenir de ces déchets. Le protocole d’intervention doit donc intégrer cette dernière phase afin de disposer de ces matières organiques de manière adaptée, éventuellement en les valorisant par compostage ou méthanisation. Dans le guide « Accompagner le traitement des déchets de plantes exotiques envahissantes issus d’interventions de gestion » édité en 2022, se trouvent quelques informations sur les méthodes de traitement d’H. verticillata mais d’autres éléments concernant des espèces proches comme Egeria densa et Lagarosiphon major aux possibilités similaires de traitement pourront également être consultés.
Commentaires
Hydrilla verticillata est sans conteste une espèce aux capacités de colonisation de nouveaux biotopes particulièrement efficaces, une « grande plasticité adaptative » selon Patrick & Singarayer (2021) dans leur revue des facteurs pouvant influencer sa répartition mondiale (facteurs géographiques, climatiques et environnementaux). Après un afflux de recherches issues des Etats Unis, conséquence très directe des difficultés de diverses natures que l’espèce y causait, le corpus d’informations s’est élargi aux autres territoires colonisés, avec en parallèle une amélioration continue des approches génétiques ayant conduit à l’identification de cinq principaux biotypes (Tippery, 2023) permettant de développer des recherches plus précises sur l’écologie et les modèles de dispersion de ces biotypes.
Avec ses premières observations sur le territoire de Guyane, ce qui est déjà connu sur l’espèce devrait nécessairement conduire à la mise en place d’une stratégie de gestion efficace. Elle devra être accompagnée d’une stratégie de communication destinée à tous les publics concernés pour que H. verticillata ne devienne pas une plante exotique envahissante largement répandue en Guyane.
Pour en savoir plus
Rédaction : Alain Dutartre (Expert Indépendant)
Relectures : Jules Mioche (FREDON Guyane), Jessica Launay (DGTM) et Clara Singh (Comité français de l’UICN)