
Si les perturbations environnementales jouent un rôle important dans la facilitation de colonisation de nouveaux milieux par des plantes exotiques, les traits biologiques de ces espèces leur permettant ces colonisations sont également un facteur explicatif majeur. Ils peuvent inclure des attributs morphologiques, chimiques, physiologiques, etc., en interactions avec les facteurs biotiques et abiotiques des biotopes dans lesquels s’installent ces espèces. De meilleures connaissances et compréhensions de ces traits et de ces interactions peuvent permettre d’améliorer les stratégies de gestion des EEE.
Parmi ces traits biologiques, la capacité de reproduction végétative à partir de fragments de tiges ou de rhizomes est largement répandue parmi les EEE, leur facilitant une dispersion rapide (voir par exemple l’article de Pyšek et Richardson 2007 sur les traits associées à l’envahissement des plantes exotiques), ce qui peut conduire à ce que certaines populations sur de vastes territoires soient en fait un seul et même clone, issues à l’origine de quelques fragments de la même plante.
La reproduction clonale
Les Jussies exotiques en métropole en sont un exemple bien connu et même si des graines fertiles leur permettant une reproduction sexuée sont maintenant régulièrement observées sur une partie croissante du territoire, la plupart de leurs colonisations ont débuté par cette seule reproduction végétative.
Les conditions de cette reproduction sont en interactions avec les facteurs environnants mais aussi directement liées aux traits biologiques du clone qui peuvent être variables au sein du genre Ludwigia. En particulier, les différents degrés de ploïdie des taxons envahissants de ce genre, c’est-à-dire les nombres de jeux complets des chromosomes présents par cellule, peuvent induire des réponses différentes des propagules aux conditions environnementales du site d’accueil.
Différentes recherches ont déjà été menées en France et ailleurs pour mesurer les capacités de régénération des Jussies à partir de morceaux de tiges, la production de biomasse et la fragmentation ultérieure des plantes étant les moteurs efficaces de leurs colonisations (voir par exemple, Dandelot, 2004 ; Dutartre et al., 2007 ; Haury et al., 2014 ; Lambert et al., 2010 ; Thouvenot et al., 2013a et b).
Après une première expérimentation à partir de fragments de pousses clonales sur deux espèces de Ludwigia invasives différant par le niveau de ploïdie (Grewell et al., 2016), Brenda Grewell et ses collègues, ayant constaté que le taxon diploïde Ludwigia peploides subsp. montevidensis montrait des capacités de développement plus importantes que le taxon décaploïde L. hexapetala, (synonyme de Ludwigia grandiflora subsp. hexapetala) ont réalisé une deuxième expérimentation pour en préciser les conditions (Grewell et al., 2019).
Ces expériences en mésocosmes ont porté sur la croissance et des réponses écophysiologiques des pousses dans des sédiments aux teneurs différentes en nutriments.
Conditions expérimentales
Les prélèvements des fragments de plantes ont été faits en début du printemps, avant la germination des bourgeons, dans deux sites proches de San Francisco (Californie) hébergeant un des deux taxons. Des segments « normalisés » des deux taxons (40 cm de longueur, comprenant 7 nœuds de bourgeons visibles et des poids frais initiaux variant d’un taxon à l’autre, mais très similaires au sein de chaque taxons (rhizomes décaploïdes, 40,8 ± 2,2 g ; rhizomes diploïdes, 13,6 ± 0,8 g) ont été plantés durant 5 semaines dans des bassins aux conditions contrôlées de lumière et de niveaux des eaux.
Des carottes de sédiments ont été collectées dans ces deux sites et dans six autres sites également colonisés par les Jussies pour évaluer les concentrations de nutriments dans les sédiments en sites naturels et établir une base pour les traitements expérimentaux pour lesquels deux niveaux de teneurs en nutriments (faible et élevé) ont été testés.
Résultats
Croissance, biomasse et allocation des nutriments
Pour les deux taxons, la biomasse totale a augmenté avec la disponibilité des éléments nutritifs des sédiments mais le taxon diploïde a montré un taux de croissance plus élevé (Figure 1A) et a produit plus de biomasse que le décaploïde L. hexapetala (Figure 1B). Les analyses de données ont montré que la ploïdie et la disponibilité des éléments nutritifs des sédiments (Figure 1C) étaient des facteurs importants de cette allocation de biomasse. La biomasse des rhizomes a augmenté chez les deux taxons en fonction de la disponibilité en nutriments dans les sédiments mais la proportion de la biomasse totale allouée à la croissance souterraine est restée faible, l’allocation des nutriments étant majoritairement portée au-dessus du sol pour développer les tiges feuillées.

Architecture végétale
Les pousses primaires produites par le décaploïde sont plus longues que celles du diploïde (Figure 1D). Les teneurs élevées en éléments nutritifs des sédiments ont largement facilité le développement en longueur des pousses principales et le nombre de branches le long de ces pousses (Figure 1E). Le nombre de branches secondaires a également été influencé par la quantité de nutriments des sédiments (Figure 1F). Alors que le nombre total de branches produites par les deux taxons était similaire, les observations et les analyses des données suggèrent que la pousse primaire comparativement longue du décaploïde était la principale différence d’architecture qui contrastait avec les branches plus denses et plus courtes du diploïde mais les différences de longueur de pousse entre les niveaux de ploïdie n’étaient pas significativement différentes.
Mesures photosynthétiques
Les deux taxons ont montré une augmentation significative du taux de photosynthèse avec l’augmentation de la disponibilité des éléments nutritifs dans les sédiments sans que le niveau de ploïdie n’induise de différence significative.
Chimie des feuilles
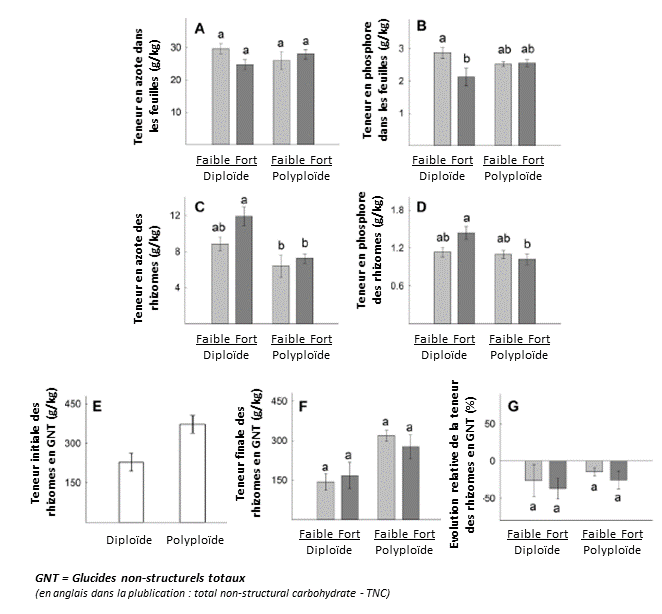
Les concentrations d’azote dans les feuilles étaient les mêmes pour les deux taxons (Figure 2A). La concentration en phosphore des feuilles était plus faible pour le diploïde avec une disponibilité accrue des éléments nutritifs dans les sédiments, mais a montré peu de changement pour le décaploïde (Figure 2B).
Réserves de stockage de carbone et de nutriments
L’azote dans les rhizomes était significativement plus élevé chez le diploïde (Figure 2C), en particulier aux concentrations élevées de nutriments dans les sédiments. Le phosphore dans les rhizomes était également plus élevé chez le diploïde (Figure 2D). Au début de l’expérience, les deux taxons avaient dans leurs fragments de rhizomes des réserves significatives de glucides non structuraux (TNC), bien que ces réserves de stockage de carbone aient été nettement plus élevées dans les rhizomes décaploïdes (Figure 2E). À la fin de l’expérience, les deux taxons avaient consommé ces réserves de carbone, mais le décaploïde avait conservé plus de réserves de stockage de carbone que le diploïde (Figures 2F et G).
Commentaires
Les résultats de ces expérimentations sur deux taxons de Jussies de niveaux différents de ploïdie montrent qu’ils ont tous deux des capacités importantes de régénération de plantes à partir de fragments de rhizomes et que les quantités de nutriments disponibles dans les sédiments sont un facteur favorable pour la production de biomasse. Ces résultats ne font que confirmer les acquis des travaux de recherches antérieurs : la reproduction végétative des espèces clonales et la régénération des plantes partir des banques de bourgeons de fragments clonaux est une stratégie de colonisation efficace dans des milieux supportant de fréquentes perturbations comme les milieux aquatiques.
Les auteurs notent sur ce point que ces capacités de régénération devraient justifier la mise en place de stratégies de gestion visant à prévenir l’érosion des berges et d’autres perturbations susceptibles de produire et de transporter des fragments de rhizome.
En revanche, la confirmation que, dans ces conditions expérimentales, le taxon diploïde présente de meilleures capacités d’absorption, d’utilisation et d’allocation des ressources au sein de la plante en développement que le décaploïde, est un résultat tout à fait intéressant.
Malgré d’importantes réserves initiales de stockage de carbone dans les rhizomes décaploïdes, le taxon diploïde a en effet montré une meilleure efficacité pouvant influencer la survie des plantes lors de leur installation, la dynamique ultérieure de ses populations et donc le succès de la colonisation.
En métropole, les observations menées depuis environ trois décennies dans différentes régions sur les dynamiques des deux taxons envahissants ne semblaient pas, encore récemment, tout à fait convergentes, citant l’un ou l’autre comme le colonisateur le plus efficace et, même dans les sites où les deux taxons coexistaient, il semblait difficile d’identifier celui qui devenait dominant, ce qui a suscité quelques débats entre différents observateurs. La diversité écologique des sites colonisés et leurs historiques très différents peuvent expliquer la récurrence de ces échanges.
En matière de gestion, dans les sites déjà fortement colonisés, cette différence d’efficacité « écologique » nous semble présenter une importance relative, car les occupations des biotopes et la production de biomasse des deux taxons sont assez similaires et peuvent donc amener à des interventions de régulation de même nature et importance. Il n’en est peut-être pas de même pour ce qui concerne les fronts de colonisation ou les installations débutant dans de nouveaux sites jusqu’alors dépourvus de ces envahisseuses et la mise en place de démarches de prévention ou d’interventions ayant plutôt des objectifs d’éradication de ces nouvelles installations localisées.
Concernant le taxon décaploïde, les auteurs de l’article notent par exemple que dans la mesure où il semble massivement investir dans l’allongement des tiges, le recours à des organismes de lutte biologique épuisant directement les réserves de stockage souterraines ou la translocation du carbone de la partie aérienne de la plante vers les organes de stockage souterrains pourrait potentiellement améliorer des stratégies de gestion intégrée. Ils citent également d’autres options telles que l’élimination mécanique des plantes ou l’utilisation d’agents de lutte biologique réduisant la biomasse des pousses.
A propos du taxon diploïde, ils indiquent que les stratégies de gestion devraient donner la priorité à une réponse rapide dans les sites nouvellement colonisés, et que la réduction des charges de nutriments dans les milieux aquatiques pourrait être une stratégie de gestion globale plus efficace pour contrôler l’établissement de ce taxon plutôt que le décaploïde.
Rédaction : Alain Dutartre, expert indépendant
Relectures : Madeleine Freudenreich, Comité français de l’UICN
Bibliographie :
Article principal : Grewell B.J., Futrell C.J., Iannucci M.T., Drenovsky R.E., 2019. Resprouting potential of rhizome fragments from invasive macrophyte reveals superior colonization ability of the diploid congener. AoB PLANTS 11: Vol. 11, No. 6, 10 p. https://doi.org/10.1093/aobpla/plz071
Publication antérieure : Grewell, B. J., Skaer Thomason, M. J., Futrell, C. J., Iannucci, M., & Drenovsky, R. E. 2016. Trait responses of invasive aquatic macrophyte congeners: colonizing diploid outperforms polyploid. AoB Plants, 8. doi:10.1093/aobpla/plw014
Pour aller plus loin :
- Dandelot S., 2004. Les Ludwigia spp. invasives du sud de la France : historique, biosystématique, biologie et écologie. Thèse de l’Université de droit, d’économie et des sciences d’Aix-Marseille. 218 pp.
- Dutartre A. (Coord.), Haury J., Dandelot S., Coudreuse J., Ruaux B., Lambert E., Le Goffe P., Menozzi M.-J., Cazaubon A., 2007. Les jussies : caractérisation des relations entre sites, populations et activités humaines. Implications pour la gestion. Rapport final – Programme de recherche Invasions Biologiques 2003-2006. Cemagref REBX Bordeaux & Ministère de l’Ecologie : 87 p. + annexes.
- Haury J., Druel A., Cabral T., Paulet Y., Bozec M., Coudreuse J., 2014. Which adaptations of some invasive Ludwigia spp. (Rosidae, Onagraceae) populations occur in contrasting hydrological conditions in Western France? Hydrobiologia, 737: 45–56. https://doi.org/10.1007/s10750-014-1815-7
- Lambert, E., Dutartre, A., Coudreuse, J., Haury, J. – 2010. Relationships between the biomas production of invasive Ludwigia species and physical properties of habitats in France. Hydrobiologia, n° 656, p. 173-186 https://doi.org/10.1007/s10750-010-0440-3
- Pyšek P, Richardson D. 2007. Traits associated with invasiveness in alien plants: where do we stand? In: Nentwig W, ed. Biological invasions. Berlin: Springer, 97–125.
- Thouvenot L., Haury J., Thiébaut G., 2013a. A success story: water primroses, aquatic plant pests. Aquatic Conservation: Marine and Freshwater Ecosystems, 23 (5) : 790-803.
- Thouvenot L, Haury J, Thiébaut G. 2013b. Seasonal plasticity of Ludwigia grandiflora under light and water depth gradients: an outdoor mesocosm experiment. Flora, 208, 430-437